Genus: Masiakasaurus Sampson, Carrano and Forster, 2001
Classification: Dinosauria - Saurischia - Theropoda - Ceratosauria - Abelisauroidea - Noasauridae
Etymology: From masiaka (Malagasy, meaning vicious), sauros (Greek, meaning lizard)
Synonyms:none
Type species: M. knopfleri Sampson, Carrano and Forster, 2001
Other Species: none
Diagnosis: as for type species
Species: Masiakasaurus knopfleri Sampson, Carrano and Forster, 2001
Etymology: knopfleri (after singer/songwriter Mark Knopfler, whose music inspired expedition crews).
Synonyms: none
Holotype:UA 8680, well-preserved right dentary with several teeth
Referred Specimens:
FMNH PR 2108-2182: maxilla; dentaries; splenial; cervical, dorsal, sacral and caudal vertebrae; dorsal rib; humeri; manual phalanges and ungual; pubes; femora; tibiae; tibia/fibula/astragalocalcaneum; metatarsals II and III; pedal phalanges and unguals.
UA 8681-8696: dentary; cervical, dorsal, and caudal vertebrae; femora; tibia/astragalocalcaneum; pedal phalanges; and unguals.
Hypodigm The holotype and referred specimens: dentaries (FMNH PR 2177-79, 2222; UA 8682); splenial (FMNH PR 2124); angular (?) (FMNH PR 2166); maxilla (FMNH PR 2183); premaxillary/anterior dentary teeth (FMNH PR 2165, 2180, 2226); posterior maxillary/dentary teeth (FMNH PR 2164, 2170, 2181–82, 2198–2201, 2220–21, 2228); cervical (FMNH PR 2139–41), dorsal (FMNH PR 2111, 2113–14,
2137–38, 2144–45, 2171, 2207, 2229; UA 8701), and sacral vertebrae (FMNH PR 2142); proximal (FMNH PR 2133, 2204, 2230), medial (FMNH PR 2110, 2125–26; UA 8688, 8692), and distal (FMNH PR 2127–28, 2156–57, 2162–63, 2168, 2202–03; UA 8689–91, 8695–96, 8702–03) caudal vertebrae; humeri (FMNH PR 2143; UA 8693–94); manual phalanges (FMNH PR 2132, 2205, 2224–25, 2227) and unguals (FMNH PR 2136, 2169); pubes (FMNH PR 2108–09); femora (FMNH PR 2115, 2117, 2120, 2123, 2148–50, 2153, 2208, 2215; UA 8681, 8684, 8712); tibiae (FMNH PR 2112, 2118–19, 2121,
2152, 2214; UA 8685, 8687, 8710–11); tibia with partial fibula and astragalocalcaneum (FMNH PR 2116, 2122); calcaneum (FMNH PR 2235); metatarsals II (FMNH PR 2147, 2151, 2154, 2175, 2206; UA 8683), III (FMNH PR 2146, 2155), and IV (FMNH PR 2214, 2234); pedal phalanges (FMNH PR 2129–31, 2136, 2158–61, 2167, 2172–74, 2176, 2216–19, 2223; UA8686, 8700, 8713–14) and unguals (FMNH PR 2134–35, 2236).
Time: Late Cretaceous (Maastrichtian)
Horizon: All specimens are from the Anembalemba Member of the Maevarano Formation
Location: Mahajanga Basin, near the village of Berivotra, northwestern Madagascar. With few exceptions, elements attributable to Masiakasaurus, including the holotype dentary (UA 8680), were
recovered as isolated specimens from a 3-m2 area in one stratigraphic horizon of a single locality, MAD 93-18. Eight additional localities (MAD 93-30, 93-35, 95-05, 99-26, 99-30, 99-38, 99- 50, and 99-51) in the Anembalemba member have produced limited materials of Masiakasaurus.
Total length: ~1.8-2 m.
Mass: 40-60 kg
Diagnosis: Differs from all known theropods in that the four dentary teeth most rostral in position are procumbent, with the first tooth set in a large, ventrally expanded alveolus that is almost
horizontal in orientation. Also differs from all known theropods in that it has a strongly heterodont lower dentition: the first four teeth are elongate and weakly serrated, with labiolingually positioned
carinae. Each of these four teeth terminates in a pointed apex that hooks caudally. The teeth become increasingly recurved and transversely compressed with increasing caudal position in the jaw, and
possess more standard, mesiodistally positioned carinae (Sampson, Carrano and Forster, 2001) .
Diagnosis (based on holotype only) Abelisauroid theropod with anterior four dentary teeth procumbent, the first inclined at 108 above the horizontal and lying in an alveolus that is slung below the ventral margin of the dentary. First alveolus large and ventrally expanded, lying lateral to an anteroposteriorly long dentary symphysis. Lower dentition markedly heterodont: the first four teeth are weakly spoon-shaped, elongate, and terminate in a posteriorly hooked, pointed apex. Anterior dentary teeth bear two weakly serrated posterior carinae and have faint posterior ridges; more posterior teeth are transversely compressed, recurved, and have a serrated posterior carina (Carrano, Sampson and Forster, 2002).
Comments: Here we report the discovery of a small-bodied (~1.8 m) predatory dinosaur from the Late Cretaceous (Maastrichtian) of Madagascar. Masiakasaurus knopfleri, gen. et sp. nov., represented
by several skull elements and much of the postcranial skeleton, is unique in being the only known theropod with a highly procumbent and distinctly heterodont lower dentition. Such a derived dental morphology is otherwise unknown among dinosaurs. Numerous skeletal characteristics indicate that Masiakasaurus is a member of Abelisauroidea, an enigmatic clade of Gondwanan theropods. Previously, small-bodied abelisauroids were known only from Argentina1±3. The occurrence of Masiakasaurus on Madagascar suggests that small-bodied abelisauroids, like their larger-bodied counterparts, were more cosmopolitan, radiating throughout much of Gondwana and paralleling the diversi®cation of small coelurosaur theropods in Laurasia (Sampson, Carrano and Forster, 2001).
Together, the specimens referred to Masiakasaurus knopfleri account for about 40% of the skeleton. The concentration of isolated Masiakasaurus elements at MAD 93-18 includes remains of at least six individuals. All of these specimens are assigned to a single species, despite many of the elements (for example, femora, tibiae and vertebrae) being represented by several specimens, as none shows evidence of belonging to more than one taxon of small-bodied non-avian theropod. Two osteological features (closure of the vertebral sutures and fusion of the crural and tarsal elements) indicate that the largest materials represent adult or near-adult individuals of this small-bodied taxon (Sampson, Carrano and Forster, 2001).
Carrano et all. described the osteology of the new small theropod dinosaur Masiakasaurus knopfleri, from the Late Cretaceous Maevarano Formation of northwestern Madagascar. Approximately 40% of the skeleton is known, including parts of the jaws, axial column, forelimb, pelvic girdle, and hind limb. The jaws of Masiakasaurus are remarkably derived, bearing a heterodont, procumbent dentition that is unknown elsewhere among dinosaurs. The vertebrae are similar to those of abelisauroids in the reduction of the neural spine, lack of pleurocoelous fossae on the centrum, and extensively pneumatized neural arch. The limb skeleton is relatively gracile and bears numerous abelisauroid synapomorphies, including a rounded humeral head, peg-and-socket iliac-pubic articulation, prominent femoral medial epicondyle, expanded tibial cnemial crest, and double-grooved pedal unguals. The femora and tibiae show evidence of dimorphism. More specific features shared between Masiakasaurus, the Argentine Noasaurus, and the Indian Laevisuchus suggest that these taxa form a clade (Noasauridae) within Abelisauroidea. This is supported by a cladistic phylogenetic analysis of 158 characters and 23 theropod taxa. Additionally, Ceratosauria is rendered paraphyletic in favor of a sister-taxon relationship between Neoceratosauria and Tetanurae that is exclusive of Coelophysoidea. The unique dental and jaw specializations of Masiakasaurus suggest deviation from the typical theropod diet. Finally, the distribution of noasaurids further supports a shared biogeographic history between South America, Madagascar, and India into the Late Cretaceous (Carrano, Sampson and Forster, 2002).
The overall morphological similarities between Masiakasaurus, Noasaurus, and Laevisuchus are difficult to evaluate becasue the latter two taxa are extremely incomplete. Nonetheless, Noasaurus and Masiakasaurus share several synapomorphies (cervical neural spine placed on the anterior half of the centrum, reduced shaft of metatarsal II, simple maxillary palatal process, and prominent rim on the antorbital fossa) that suggest these two taxa form a clade, Noasauridae. Laevisuchus shares features with abelisauroids (Novas and Bandyopadhyay, 1999) and noasaurids (anterior placement of cervical neural spine, cervical epipophyses small posteriorly). Additional specimens will permit a more detailed analysis of the relationships of these three theropods.
Features throughout the skull and skeleton reveal Masiakasaurus to be an abelisauroid theropod. More specifically, it may belong to the clade Noasauridae, containing the Late Cretaceous Noasaurus, from
South America, and Laevisuchus, from India. If so, the geographic distribution of this clade parallels that of carnotaurine abelisaurids, further supporting a hypothesis of shared biogeographic history for South America, Madagascar, and India. Finally, the presence of Masiakasaurus on Madagascar documents a more global distribution of small-bodied abelisauroids than has been previously appreciated. (Carrano, Sampson and Forster, 2002).
More puzzling still is the fact that the posterior dentary and maxillary teeth of Masiakasaurus retain the more generalized carnivorous morphology seen in most other theropods. Thus, unlike therizinosaurs, the entire dentition has not been modified from the basic theropod plan; changes are localized to the anterior region. The posterior teeth are recurved, laterally compressed, and finely serrated. In addition, the serrated carinae are anteriorly and posteriorly positioned, suggesting that the teeth were used for holding, cutting, and/or slicing animal tissues. The combination of this morphology with that of the anterior dentition is not yet explicable. One possibility is that Masiakasaurus was insectivorous or piscivorous, using its anterior teeth for acquiring small, whole prey items and its posterior teeth for maceration. More complete materials of the skull and forelimb will undoubtedly shed light on the feeding and predatory habits of Masiakasaurus (Carrano, Sampson and Forster, 2002).
As might be expected with any sample including multiple individuals, marked variation exists among the numerous specimens of Masiakasaurus. Specifically, two discrete morphs are present in the sample and are demonstrable on comparably sized elements. These ‘‘gracile’’ and ‘‘robust’’ morphs are most easily discerned on the appendicular elements, probably because these have the largest sample sizes. The ‘‘robust’’ morph is characterized by pronounced muscular and ligamentous attachments, whereas these sites are less distinct in the ‘‘gracile’’ morph. In addition, ‘‘robust’’ tibiae show fusion with the tarsal
elements, whereas this fusion is lacking in ‘‘gracile’’ tibiae (Carrano, Sampson and Forster, 2002).
Illustrations:
Masiakasaurus knopfleriholotype dentary (UA 8680) in right lateral view.

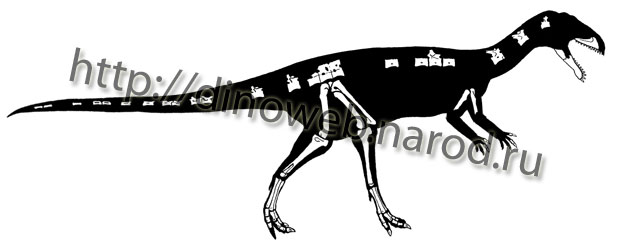
Composite skeletal reconstruction of Masiakasaurus knopfleri in left lateral view, based on the holotype and referred materials. The materials recovered to date represent approximately 40% of the skeleton. Approximate length of adult skeleton is estimated at 1.8 m.
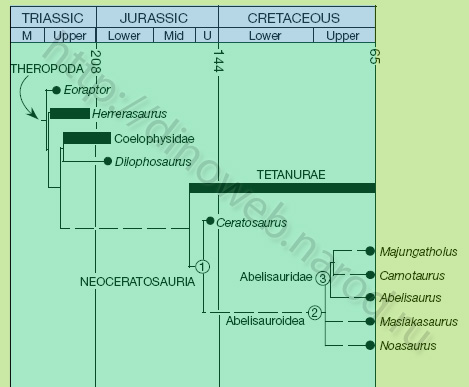
Stratigraphically calibrated cladogram of phylogenetic relationships of Abelisauroidea, including Masiakasaurus. Unambiguous synapomorphies supporting the nodes are: (1) (Neoceratosauria), lateral temporal fenestra much larger than orbit; numerous foramina surrounding axial diapophysis; notched posterior iliac margin; pegand-socket articulation between ilium and pubis; femoral fourth trochanter reduced to low ridge; deeply excavated medial surface of proximal fibula; enlarged ilio®bularis tubercle on fibular shaft; (2) (Abelisauroidea): caudal dentary margin with notch for surangular; enlarged external mandibular foramen; cervical neural spine abbreviated craniocaudally; seven sacral veretebrae; large rounded coracoid; humerus straight-shafted with bulbous head; hypertrophied mediodistal crest on femur; large tibial cnemial crest with laterallydirected hook; distal; cnemial crest curved dorsally; rectangular astragalar ascending process; triangular set of grooves on lateral and medial pedal unguals; and (3) (Abelisauridae), craniofacial elements with external sculpturing; long, shelf-like maxilla-jugal contact; greatly exposed paradental plates bearing a series of vertical ridges and grooves; dorsal veretebral parapophyses projecting far laterally from centrum; distal ends of caudal transverse processes expanded craniocaudally; forelimb short relative to other skeletal elements. M, Middle; U, Upper. Dates are millions of years before present (modified from Sampson, Carrano and Forster, 2001).
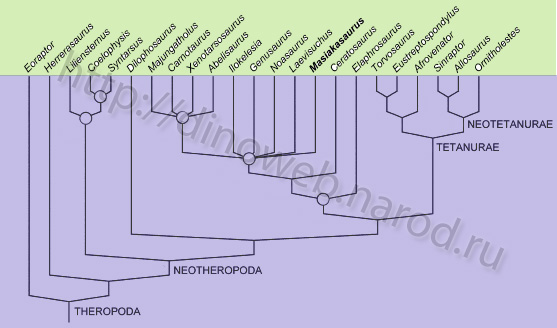
Cladogram showing phylogenetic relationships of Masiakasaurus knopfleri and other theropods (modified from Sampson, Carrano and Forster, 2001).
References:
Sampson, S. D., M. T. Carrano, and C. A. Forster. 2001. A bizarre new predatory dinosaur from Madagascar. Nature 409: 504–506.
Carrano, Sampson and Forster, 2002. The osteology of Masiakasaurus knopfleri, a small abelisauroid (Dinosauria: Theropoda) from the Late Cretaceous of Madagascar. JVP 22(3) 510-534.
© Tetrapoda Database Roman Ulansky roman.ulansky@gmail.com or adios85@mail.ru
|

